|
|
|
Ergebnisse
Charakterisierung hitzeinduzierter Proteine (HSP 70) durch nichtradioaktiver Nachweismethoden
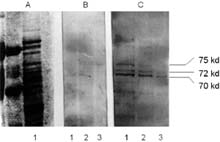
Die Versuche in Fig. 1 sollten die Bedingungen für die Stressinduktion der HSP70 Proteine bei T. thermophila festlegen. Dazu wurden die Zellen, anders als bei [5] mit 36°C, mit 34°C für 15 min. thermal gestreßt. Man erkennt eine deutliche Zunahme der HSP70 Proteine bei den induzierten Zellen (Fig.1; C I-3). Eine konstitutive Expression der HSP70 Proteine ist auch bei einer Erhöhung der Gesamtprotein-konzentration bis 100µg aus physiologisch wachsenden Zellen nach der Westernblot Analyse nicht erkennbar.
PCR Versuche mit heterologen Primern
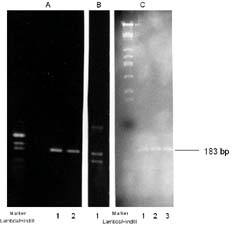
Für die Untersuchung der Homologie der konservierten Hitzeschock-gene wurden PCR Versuche mit den hsp70 bzw. 17S rDNA Primer durchgeführt. Wie in Fig.2; lane 1 und 3 erkennbar ist, läßt sich mit stringenten Primern aus dem hsp70 Gen ein PCR-Fragment aus T. thermophila DNA amplifizieren, das mit 187bp die gleiche Größe wie das PCR-Kontroll Fragment aus S. cerevisiae besitzt. Für die Überprüfung auf Hefe-Kontamination wurden dann im folgenden mehrere Kontrollen durchgeführt. So ist auf Fig. 2; lane 4 und 6 die PCR mit DNA und den hsp70-bzw. 17S rDNA-Primern (850 bp) aufgetragen worden. Der Unterschied der beiden lanes besteht lediglich in der Auftragskonzentrationen. Da in der lane 5 kein PCR-Fragment amplifiziert wurde läßt sich resümieren, daß eine Kontamination der T. thermophila-DNA sowohl während der Nukleinsäuren-Präparation, als auch bei der PCR-Durchführung, mit Hefe-DNA auszuschließen ist.
Überprüfung der Sequenzunterschiede der PCR- Produkte durch ein 1,5%iges HA-Y Agarosegel [20]
Wie bereits in [13] beschrieben, können mit dieser Methode, die hier in Fig.3 verwendet wurde, PCR-Fragmente mit gleicher Basenanzahl aufgrund ihrer Basenzusammensetzung getrennt werden. Die in Fig.2; lane 1 und 3 amplifizierten Amplikons wurden nach Elution einer Elektrophorese mit dem Farbstoff HA-Y unterzogen. Auch unter diesen Bedingungen wurden die aufgetragenen Fragmente (Fig.3 Abb. C-1 lane 1,2 und 3) nicht voneinander getrennt, während die Trennung der Mixtur bestehend aus Huhn, Ziege und Reh PCR-Fragmenten (Abb.B; lane 1) deutlich erkennbar ist. Daraus lassen sich erste Erkenntnisse bezüglich einer starken Homologie der Basenzusammensetzung der Hefe und T. therrmophila PCR-Fragmente ziehen.
Northernblot Analyse
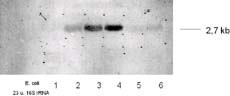
Die folgenden Northernblot Analysen sollten einerseits die gewonnene homologe T. thermophila hsp70 Sonde überprüfen und andererseits den hsp70 mRNA-Gehalt nach unterschiedlichen Hitzeschockzeiten darstellen. Die nichtradioaktiven Northernblot Analysen ergaben eine Induktionskurve der mRNA mit einer Größe von 2,7kb (Fig.4; Abb.A/B lane 1-4,), die ihr Maximum nach 10 min. thermalen Stresses erreichte. Bereits 5 bzw. 10 min. (lane 5 und 6) später ging der mRNA-Gehalt bereits auf das Niveau der unbehandelten Kontroll mRNA zurück. Die Ergebnisse dieser Analysen führen zum Schluß, daß es sich hierbei um das induzierbare Hitzeschockgen hsp70 von Tetrahymena thermophila handeln muß. |
|
|
|
|
|
Material und Methoden
Chemikalien
Trypton und Pepton wurde von Gibco, Felll-EDTA von Sigma bezogen. Alle Detektionsreagenzien (ELISA, Nichtradioaktive Hybridisierungsreagenzien) erhielten wir von Boehringer Mannheim bzw. Promega. Die Primer wurden von MWG bezogen.
Zellkultivierung und Stressinduktion
1 Liter Medium nach [ 6 ] wurde autoklaviert und Tetrahymena thermophila Zellen (Stamm 5) bis zu einer Dichte von 1-2x10' wachsen gelassen. Die sich in der log-Phase befindlichen Zellen wurden aliquotiert und bei 34°C mit unterschiedlichen Zeiten hitzeinduziert (siehe Northernblot-Legende).
Gesamtprotein-Extraktion und PAGE
T. thermophila Zellen wurden 5min. mit 8000xg zentrifugiert und das Pellet in 500µl Lysispuffer wie in [7] beschrieben lysiert. Der Überstand wurde verworfen und 10, 50 und 100µl Proteinlysat vor dem Beladen auf ein Polyacrylamidgel [8] drei Minuten in Probenpuffer [9] durch Kochen denaturiert.
Westernblot Analyse und Detektion
Nach der Elektrophorese wurden die Proteine in Transferpuffer [9] bei 200 V in 90 min. mittels einer Elektrodenkammer auf Nitrocellulose (Hybond C-extra, Amersham) transferiert. Nach dem Transfer wurde die Nitrocellulose nach dem Protokoll der Firma Promega behandelt und mit dem humanen monoklonalen Antikörper der Firma Dianova inkubiert. |
|
|
|
|
|
|
Diskussion
Diese Arbeit zeigt im ersten Teil die Erhöhung des Gehaltes der HSP70 Proteine . Zu sehen sind auf Fig.1 (Abb.C lane 5, 6 und 7) drei Banden nach dem Immunoblotten und dem Immunostaining mit dem humanen monoklonalen Antikörper nach Hitzeschock. Hierbei handelt es vermutlich um Isoformen des gleichen HSP70 Proteins nach Denaturierung mit Natrium dodecyl sulfat [15]. Avides Carmo, M. et al. konstruierten ein Oligonukleotid aus zwei HSE Regionen und führten damit ein Bandshift Assay durch [16]. Auch hier wurde zur Analyse der Promotorregion neben einem Drosophila-HSE ein Hefe-HSE verwendet. Anders als bei dem 33 mer Oligomer von Amaral, M. D. et al., das zur Hybridisierung benutzt wurde, welches aus der Aminosäurensequenz mehrerer Organismen [17] unter Berück-sichtigung der Codon Usage von T. pyriformis konstruiert wurde, wurde in dieser Arbeit absichtlich mit stringenten Primem gearbeitet.
Trotz der beobachteten unterschiedlichen Codon usage [18,19] wurden Primer nach Abgleich aus der EMBL-Datenbank aus dem homologen codierenden Bereich des hsp70 Gens von Saccharomyces cerevisiae synthetisiert und direkt in die PCR eingesetzt. Erste Hinweise über die Homolgie der zuvor amplifizierten PCR-Fragmente zeigten dann auch die Versuche mit dem Farbstoff HA-Y. Die Sequenzierung der S. cerevisiae- und T. thermophila-Amplikons ergab dann auch eine 100%ige Homologie. Ein weiterer Schluß läßt sich aus den geschilderten Ergebnissen ziehen. Das Codon-usage, so wie es von Martindale, D. W. beschrieben wurde, trifft zumindest bei dem untersuchten Hitzeschockgen hsp70 von T. thermophila und im Vergleich S. cerevisiae nicht zu.
Die (in Fig. 4 lane 1, 5 und 6) abgebildeten Northernblot Analysen bestätigen die bereits von Amaral, M. D. et al. bei T. pyrifomis gemachten Beobachtungen, daß die mRNA für das hsp70 Gen nur in hitzeinduzierten präsent ist, nicht aber in physiologisch wachsenden Zellen. Dieses Phänomen läßt sich mit den bereits von Amaral, M. D. erzielten Ergebnissen erklären. Demnach scheinen auch bei T. thermophila die induzierten HSP70 Proteine einer negativen Feedback-Regulation zu unterliegen. Sämtliche bisherigen Versuche, die sich auf die Analyse beziehungsweise Untersuchung des hsp70 Gens bei T. thermophila oder T. pyriformis beziehen, wurden mit heterologen Sonden durchgeführt.
In dieser Arbeit konnte gezeigt werden, daß es für die Untersuchung und Isolation des hsp70 Gens keinerlei Modifikation der DNA-Sonden bedarf. |
|
|
Literatur
1. Goldberg, C. J., Y. Luo, M. Fenna, R. Baler, R. Weinmann and R. Voellmy. 1988. Purified human factor activates heat shock promotor in HeLa cell-free transcription system. J. Biol. Chem. 263:19734-19739.
2. Sorger, P. and H. R. B. Pelham. 1987. Purification and characterization of a heat shock element binding protein from yeast. EMBO J. 6:3035-3041.
3. Weiderrecht, G., D. Shuey, W. Kibbe and C. Parker. 1987. The Saccharomyces and Drosophila heat shock transcription factors are identical in size and DNA binding properties. Cell 48:507-515.
4. WU, C., S. Wilson, B. Walker, I. Dawid, T. Paisley, V. Zimarino and H. Ueda. 1987. Purification and Properties of Drosophila heat shock activator protein. Science 238:1247-1253.
5. Amaral, M. D., l. Galego and C. Rodrigues-Pousada. 1988. Stress response of Tetrahymena pyriformis to arsenite and heat shock : Differences and similarities. Eur. J. Biochem. 171:463-70.
6. Yu , G. L., m. Hasson and H. Blackburn. 1988. Circular ribosomal DNA plasmids transform Tetrahymena thermophila by homologous recombination with endogenous macronuclear ribosomal DNA. Proc. Acad. Sci. USA. 85:5151.
7. Danielian, S., R Fagard, I. Boulet and S. Fischer 1990. The retarded electrophoretic migration of p56Ick induced by vanadate in lymphoma cells correlates with modified kinase activity. Biochemie 72:315-322.
8. Laemmli, H. K. 1970. Cleavage of structural proteins during the assembly of the head of bacteriphage T4. 227:680-685.
9. Manatis, T., E. F. Fritsch and J. Sambrook. 1989. Molecular cloning. Sec. Edition, Cold Spring Harbor Lab. Press, New York.
10. Corr, R and W. Eichler. 1985. Isolation and Properties of Macronuclei from Tetrahymena Thermophila. Protozol. 32:45-49.
11. Pharmacia protokoll Macropreparation kit for genomic DNA
12. Saiki, R. K., D. H. Gelfand, S. Stoffel, S. J. Scharf, R. Hiuchi, G. T. Horn, K. B. Mullis, H. A. Erlich. 1988. Primer-directed enzymatic amplification of DNA with a thermostable DNA Polymerase. Science 239:487-491.
13. Waver, C.; H. Rüggeberg; G. Meyer; G. Muyzer. 1995. A simple and rapid electrophoresis method to detect sequence variation in PCR-amplified DNA fragments. Nucl. Acid. Res. 23:4928-4929.
14. Gene screne Manual (Du Pont)
15. Margulis, B. A., P. V. Nacharov, O. I. Tsvetkova, M. Welsh and A. V. Kinev. 1991. The characterization and use of different antibodies against the hsp70 major heat shock protein family for the development of an immunoassay. 12:670-673.
16. Carmo do, M., C. E. Sunkel, P. Mordas-Ferreira and C. Rodrigues-Pousada. 1990. Properties and partial characterization of the heat-shock factor from Tetrahymena pyriformis. Eur. J. Biochem. 194:331-336.
17. Amaral, M. D., L. Galego and C. Rodrigues-Pousada. 1993. Heat-shock-induced protein synthesis is responsible for switch-off of hsp70 transcription in Tetrahymena. Biochemica et Biophysica Acta. 1174:133-142.
18. Martindale, D. W. 1989. Codon Usage in Tetrahymena and other Ciliates. J. Protozool. 36(1):29-34.
19. Horowitz, S. and M. A. Gorovsky. 1985. An unusual genetic code in nuclear genes of Tetrahymena. Proc. Natl. Acad. Sci. 82:2452-2455.
20. Hanse Analytik GmbH, Fahrenheitstraße 1 (BITZ), 28359 Bremen, Tel.: 0421-2208196, Fax:0421-2208278. |
|
|